

Ordained-Becoming (Part 2)
(….please read the previous post first.)
The Positive Constraints of Epigenetics
When scientists first compiled the genetic sequences of wildly diverse organisms they were stunned and confused by a very unexpected discovery. Organisms as starkly different as a human and a fruit fly shared many similar gene clusters. Two organisms, of vastly different scale, with vastly different niches, and vastly different heritages seemed to be made of the same code. No biologist was expecting this. The conventional expectation was that humans were wired with distinctive and specialized genes. Everyone certainly expected humans to contain a lot more genetic information than the miniscule fruit fly. It was inconceivable humans would have roughly the same number of genes (29,000) as flies. Where, for instance, was that vast instructional manual for building a brain? Ernst Mayr, the grandfather of modern evolutionary synthesis, once declared “the search for homologous genes is quite futile except in very close relatives.” But now modern genetic sequencing shows that homologous genes are everywhere. Despite the apparent difference in complexity between the smidgen of a fruit fly and human with its powerful mind, both organisms are built from the same set of “standard” informational blocks. The development of animal eyes, limbs, heads and hearts, though obviously different in every phyla from fly to reptile to fish, and long believed to have evolved independently, are all governed by the same genes. A common set of bits to code us all, at least among animals. Humans and chimps, for instance, share so many identical genes, that by the normal statistical measure of 99%, we should be declared identical species.
What’s going on? According to modern Darwinian theory every gene on every chromosome should be under constant, daily scrutiny by evolutionary pressure, and should therefore be in constant churning flux. The genotypes of organisms should reflect the diversity of their phenotypes. Primitive flies without gigantic brains, pancreas, and five fingers like us should not have our genes, nor we theirs! Yet, the new tools show that genes are conserved faithfully, stubbornly. Many – if not most – of our genes were cast hundreds of millions of years ago, long before anything like a human walked on land. They should not be called human genes. While the continents shifted, poles reversed, extreme climates came and went, and orders of creatures appeared and disappeared, complexes of genes persisted unchanged for millennia. As evolutionary geneticist Sean Carroll notes, the basic “tool kit” of genes spawning the myriad bilateral creatures which burst forth in the Cambrian, predated the Cambrian, “the mammalian tool kit predated the rapid diversification of mammals in the Tertiary period, and the human tool kit long predated apes and other primates.”
We are built from ancient potential. Evolutionary potential, in the form of stable ancestral gene kits, precedes the particular genetic arrangement of a species. It requires the long hard scrubbing of natural selection to move organisms into the potential opened by the genetic tool kits. As Carroll continues in his remarks, “It is clear that genes per se were not drivers of evolution. The genetic tool kit represents possibility – realization of its potential is ecologically driven.”
The basic kit of evolution is not a gene, but a “homeobox,” which is a cluster of about 180 genes. (The genes in a homeobox cluster are abbreviated to “hox” genes.) These are the bricks. The creative power of the dual homeobox processes of evolution and development (called evo-devo) resides in the 3% of genes which regulate the hox genes. As a single fertilized egg develops into a billion diverse cells, the intricate web of switching genes activate and deactivate the hox genes, so that the proper sequences of universal bricks are powered up to make the proteins to make the tissue to make the organism. In response to changes in the environment, natural selection will deactivate (without deleting) homeoboxes, or re-order their sequence, or shift the whole cluster to a new place in line, or rewired the web of their controlling routines. Re-arranging the boxes by means of switches has no effect on the function of box (maintaining continuity), and no effect on the durability of other switches, so this modular system is incredibly robust. Carroll who authored a popular book on evo-devo, Endless Forms Most Beautiful, says “the key to the making of ‘endless’ forms (i.e. diversity) is in the astronomical number of possible combination of regulatory inputs and switches” among the homeoboxes.
Over time evolution graduates to evolving larger “chunks” which is the main way it can navigate large possibilities spaces. If monkeys can only type letters, it takes forever before they randomly type out Hamlet; but if their typewriters type of only full words, the wait is much less. If they type full sentences, it’s quicker still. Homeoboxes are like typing whole random intact paragraphs. Pretty soon you’ll have a story of some sort. In computer terms this strategy is the equivalent of evolving whole subroutines instead of merely mutating words in a program.
However the effect of evolving life via chunks of code tightly linked into webs is that these webs produce both negative constraints due to their embodiment, and positive constraints due to emergent patterns. I want to address both consequences here.
Every book, even one created by random mixes, carries within its pages a different long message. The text is its intangible code, and the ink stamped on sheaves of bound paper is the code’s tangible embodiment. But as media theorist Marshall McLuhan noticed, the form of the book itself – the medium – carries its own message regardless of the book’s content. Other media do the same. The alphabet and its literacy changed how we think, independent of what we read. Regardless of what is being shown on television, the physical display of scanned raster lines on a screen, the repeated interruptions of 30-second commercials, the rapid sequence of frames, and all the semiotic conventions of a TV show also carry a signal, sometimes one that is more powerful than its mere particular content. In McLuhan’s words, the medium itself (alphabet, book, TV, blog, twitter) is the message.
Genetic information is transmitted, like a ghost, from generation to generation. However, like all intangible information, genetic information must be embodied in tangible atoms to be transmitted and expressed. Those atoms of informational embodiment — whether electronic blips or chemical signals — constrain what can be said and often add an implicit meaning. In other words the medium itself carries a message.
The medium for genetic information is an extremely complex set of molecules, which operates at many levels of embodiment as base pairs, codons, genes, homeoboxes, chromosomes, and genome. At each structural level there is a tangled web of interactions, and multiple pathways. Consider for instance that all the specialized cells in your body, from heart cell to brain cell, have a very different biochemical form but are run from the same DNA “blueprint.” So the same blueprint can build more than one kind of building. A single gene can code more than one protein, and one protein can fold more than one way. The critical additional information for determining what kind of cell to develop at each juncture is often carried implicitly in the physical organization of the gene. The order by which genes appear on the chromosome make a difference, even though this order may not be prescribed in the genetic information itself. This order influences the ability of some genes to inhibit or active other genes, which is how a general fertilized egg cell differentiates during morphogenesis into a vast multi-varied cellular creature. The developmental path from pure DNA code to viable holistic body is a long, complicated, interacting, self-referential circuit. This web of interconnecting self-regulating genes and developmental paths is its own complex adaptive system. In some ways it is as complex as evolution itself. So, in addition to the specific code carried by particular DNA sequences, then, there is another level of messaging that is carried by the physical network of DNA, genes and chromosomes. This message-of-the-medium is called epigenetic.
Epigenetics entails all the influences genes exert outside of the information they carry within DNA. Examples of this dynamic include paramutation, wherein the physical presence of an allele (counterpart gene) can influence development without changing the heritable information. Or a gene may be silenced when its position on the chromosome is physically shifted even a small amount. In insects, sex is often determined by silencing the entire paternal genome. There are types of cell memory wherein a cell “bookmarks” genes before cell division, and the bookmarks are passed on to daughter cells, thus carrying gene expression instructions outside of the DNA. These are simple cases, but epigenetic regulatory circuits yield deep complexity in the style of “this gene inhibits that gene, except when those genes are activated by these genes regulated by that first gene.” This reflexive web of links sprouts emergent patterns that are beyond our understanding at the moment, even though their effects are real and visible. In short, the epigenome system can shift, bend, and create potential forms, while the underlying genome remains unchanged.
Homeoboxes, which may be hundreds of million years old, are like train tracks “in the tree of life”. At the fork where each new species divides the line, the track keeps running the durable “trains” of commonly shared homeobox genes in parallel. As evolution keeps diverging, constant parallel tracks of identical hox genes keep organisms returning to similar and seemingly universal solutions. Biologists call this surprising constancy “parallelism” because the genetic structures are propagating and enduring in parallel.
Therefore one way in which epigenetic structures promote convergent evolution is by conveying the same enduring parallel solutions contained in primeval hox genes. Legs and limbs, antennas and horns, snouts and noses, eyes and compound eyes keep reappearing because the primeval “tool kit” for each form has been retained deep in operating system. Combinations of these lego-like units are recombined in a mash-up fashion, or selectively suppressed, to produce innovative expressions of familiar forms.
But what conserves the genetic bundles? Why are homeoboxes so inexplicably durable despite evolution’s ceaseless adaptive scrubbing? Genetic homeoboxes remain unchanged for hundreds of millions of years primarily because their genes are arranged in networks. They are like miniature internets, with genes linking to many other genes. Some regulatory genes turn on (or off) other genes, which in turn may regulate yet others, so the entire web of interacting genes forms an extremely complex genetic ecology that is self-sustaining, and often self-repairing. We know from both experiments in the lab and from computer simulations that complex networks of self-regulating nodes – such as genomes – generate internal dynamics and “order for free.” That is, even randomly wired-up networks will produce the same recurring order – certain behaviors or forms – no matter how the nodes are arranged. In other words, types of emergent order are inevitable for that kind of network, and this order, or structure, will be produced outside of, or on top of, the structure produced by adaptive evolution.
These recurring emergent forms are much like the self-sustaining tornado that appears in draining water under proper conditions. As long as the suitable conditions continue, the inherent form is sustainable and available to be appropriated by natural selection. The emergent attractor pushes/pulls evolution to work down its path, and in this way acts as a positive constraint.
The ability of self-regulating networks to manifest emergent structures is well known and can be seen in many types of systems. The pioneering discoverer of this effect is the theoretical biologist Stuart Kauffman, who wrote the classic tome “The Origins of Order.” Inspired in part by Kauffman’s insight I also wrote a big book about how “out of control” decentralized systems can generate new levels of useful order. Complex systems as diverse as galaxies, human societies, brains, computer networks, and artificial evolution all display the ability to generate emergent, self-sustaining structures where none existed before. Kauffman brought a mathematical rigor to these intuitive hunches and proved brilliantly that these intensely networked systems – such as genomes – would inevitably generate recurring forms. Just as important, Kauffman and others also proved in theory and practice that these emergent recurring forms tended to hover at maximal evolvibility and flexibility. Which is why the homeobox clusters selected by genetic networks are primed to keep the organism evolving even while they keep its genome intact.
The key insight gained by the last three decades of research on complex adaptive systems is that the variation presented to natural selection is not random. Random mutations are often not random; variation is constrained by geometry and physics (see above); and most importantly, variations are often shaped by the possibilities inherent in the complex system (a la vortex). Once upon a time this was heresy, but as more and more biologists run computer models, the idea that variation is not random has just about become a scientific consensus.
Epigenetic networks regulating themselves tend to dictate what genes cannot do, but the same networks also favor certain kinds of complexes which can be built. This flips the traditional view. In the classical theory of evolution, an internal source (mutation) generates variation which natural selection then steers in random (but adaptive) directions. The new view is that the engine of external natural selection pushes change forward, but internal sources channel the not-completely random variation. In the old, the internal created, while the external steered; in the new, the external creates, while the internal steers. And when the internal directs, it re-directs. As early paleontologist W.B. Scott put it, epigenetic networks and the complexity of evolution create “Inherited channels for preferred change.”
In the textbook version of evolution, evolution is a mighty force propelled by a single, simple, near-mathematical mechanism: random mutations selected by adaptive survival. But the two new widely-adopted biological tools I’ve mentioned above – cheap genetic probes and strong computer models – reveal two additional legs for evolution’s power. We can now see that the creative engine of evolution stands on three legs: the adaptive, the contingent, and the inevitable.
The Tripod of Evolution
The adaptive force is good ol’ natural selection delivered by an organism’s environment. Not every convergence is driven by the internal push of constraints. Often times two isolated lineages of organisms in similar environments will re-invent solutions for the familiar reason that survival steadily requires the selection of similar adaptations. Forms converge because their environments converge.
For instance, the defensive venomous sting has been evolved 10 times: in the spider, the stingray, the stinging nettle, the centipede, the stone fish, the sea anemone, the male platypus, the scorpion, the cone shell mollusk, and the snake. A close examination in four very different animal phyla reveals that the poisons from all four disrupt the potassium channels in the neurons of the injected attacker, making a tidy case of convergence. However, the potassium pathway is apparently fairly susceptible to disruption because the four different groups were able to devise four different chemical hacks to block it. Their distinctive molecular makeup and unrelatedness make it clear each lineage evolved the hack separately, but it is not clear what they are converging upon except the vulnerability of the potassium channel.
For many evolutionists, this is all the explanation needed. Adaptation by natural selection can explain everything. But as Niles Eldridge once observed, “Any theory that explains everything, explains nothing.”
But as I believe new tools and insight have demonstrated, there are other dimensions of evolution at work beside the adaptive nature of the environment. One of those is the role of contingency – pure luck. Gould’s “Wonderful Life” is the most persuasive defense of this evolutionary factor. As he elegantly argued, a lot of what happens in evolution comes down to the lottery. A species, or even family of species, can be extinguished by drastic climate change, tectonic drift, asteroid impacts, or other “acts of God.” Whether these extinctions are a result of “bad luck” or “bad genes” is always a debate, but at smaller scale, certainly much of the fine detail of speciation is a result of contingency. That is, rewind life’s tape and it plays out different. No one argues that the individual spots on a Monarch butterfly are inevitable, or that humans had to have 32 teeth. Contingency and random luck operate in evolution; the question is, how deep into the course of evolution does it go?
The third leg of evolution’s tripod is structural inevitability. Whereas contingency can be thought of as a “historical” force, that is, a phenomenon where history matters, the structural component of evolution’s engine can be thought of as “ahistorical” in that it produces change independent of history. Run it again, and you get the same story. This aspect of evolution pushes inevitabilities.
The iconic image of evolution is the ever-branching tree. From the base of the trunk of life, heavy arms fork up into multiple lighter branches of new divergent species, and these branches divide again into multiple twigs, each of which further diverges into yet tiny twiglets, and so on, the entire tree reticulating in infinite fractal division. The picture-book theme of evolution is ceaseless divergence. Yet the third structural corner of evolution, made up of convergence, parallelism, epigenetic, material constraint, and emergent order is non-diverging. This internally driven order plays a counterforce to the relentless diverging energy of natural selection. It gathers, re-enforces, re-turns, re-runs, and moves along an inherent path.
Here is another way to explain it. Adaptive natural selection excels at supremely optimizing a form to a constantly shifting niche. That adaptive process is always very specific, very local, and very contingent on tiny historical details and chance. But adaptive optimization presents an ancient conundrum to a species: if they perfect themselves for where they are at present, they can get stuck if the environment shifts quickly – which over the span of geological time is certain to happen “frequently.” Ideally, a species should seek a balance between optimization of the present and flexibility for the future. Yet, by definition natural selection works only in the present and cannot anticipate the future. The forces behind convergence and emergence, however, keep species near optimal evolvibility, rather than optimal adaptation, and occasionally skip across optimization (good enough is better). Converging on emergent forms, remixing durable ancient subroutines, resisting over-optimization, can keep species primed for the future.
Charted, the tripod of evolution might look like this. Classical neo-dawinism diagrammed evolution as a single force:
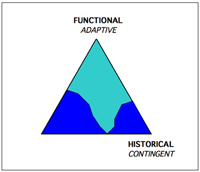
Stephen Gould emphasized contingency, once a neglected corner, as a vital force in evolution to be reckoned with, particularly on the scales of eons.
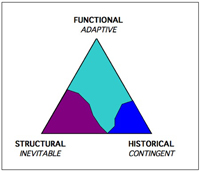
Kauffman, Conway Morris and a few other heretics reconsider the spectrum of evolution and show that it contains a third axis of structural constraints (both negative and positive) leading to the unfashionable idea of inevitabilities.
All three dynamics are present in varying proportions at different levels, counterbalancing and offsetting each other. Without the converging nature of structural emergence, it is hard to see how evolution proceeds as it has over the long term. The third dynamic can help explain how major transitions in evolution are achieved (via inherent forms ready to be exploited), and why evolution seems to have an arrow.
A metaphor comes to mind: Triple forces mold the landscape of evolution. The structural dynamics of internal constraints and emergent order carve out a deep river valley in the metaphorical landscape through which a river (adaptive selection) can meander opportunistically, but within bounds. The adaptive pressure of evolution pushes the water (evolving species) forward, but the gravity of genes keep the river within its banks at the large scale, and the sands of convergence give the meanders their distinctive universal S-shape. The detailed “particularness” of that river, all the fine contours along the shore and bottom, comes from contingency (never to be repeated), but the universal “riverness” of the river (recurring in all rivers) comes from the gravity of convergence and emergent order.
Dinosaur-ness may be a metaphorical river. Six separate dinosaur lineages have followed the same morphological pathway in evolution. Over time each of the six dino lineages displayed a reduction in their side toes, an elongation of the long bones in their paws, and a shortening of their “fingers.” Bob Bakker, the model for the dino guy in Jurassic Park, and real-life dinosaur expert claims, “this striking case of iterative parallelism and convergence … is a powerful argument that observed long-term changes in the fossil record are the result of directional natural selection, not random walk through genetic drift.”
Way back in 1897, paleontologist Henry Osborn, an early dinosaur and mammal expert, wrote: “My study of teeth in a great many phyla of Mammalia in past times has convinced me that there are fundamental predispositions to vary in certain directions; that the evolution of teeth is marked out beforehand by hereditary influences which extend back hundreds of thousands of years.”
It is important to outline what is “marked out beforehand.” In most cases, the details are contingent. The river of evolution determines the broadest outlines of form only. One might think of these as archetypes: tetrapods, snake form, eyeballs, coiled gut, egg sacks, flapping wings, repeating segmented body, trees, puffball, finger. These are general silhouettes, not individuals. Like other recurring archetypes they are patterns your brain perceives without you even noticing it – “oh, it’s a clam” it says to itself, letting you determine the particulars. On the other hand adaptive selection breeds the noticeable patterns, the flashy flourishes, the idiosyncratic specifics that belong to individuals and individual species. The weird stuff. Convergence generates the boring stuff.
The Cosmic Imperative
Baked into the very nature of nature are constraints that shape life and its unfolding in evolution. In a very indirect, yet very real, way the foundational laws of physics determine what kind of fish can swim in the oceans, or the form of animals in the tundra. The only uncertainty is how much of fishy-ness is predetermined? Is it merely their streamlined shape, air bladder, and fins (all which have evolved more than once), or does the determinacy reach down to their egg-laying, gills, and schooling behavior? Or even further to the concentric rows of teeth in a shark’s and the electrical field of the eel (which has evolved more than once)? Once unleashed, life invades all domains, never retreats, and so a water planet with life will have “fish” of some sort. Their shape will be guided in part by geometry, physics, and structural constraints. Those are constant (perhaps on other worlds, too.) The laws of physics also dictate the character of oceans themselves, and of tundras and meadow (and even whether there are oceans, tundras, and meadow), so more of life is dictated by the initial conditions of the universe than we normally appreciate.
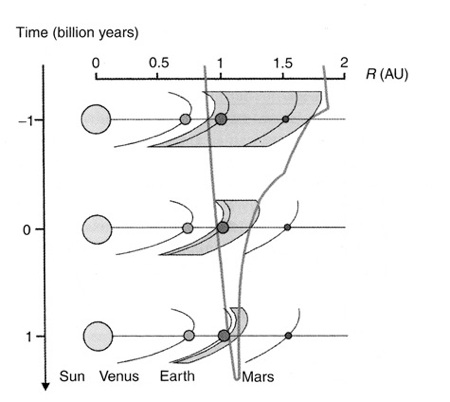
How the goldilocks zone (gray color) of Earth is narrowing over cosmological time.
That fact has led a few to examine the initial conditions of the universe to see whether it too has a tilt towards life, or particular forms of life. There are many indications the universe is biased to life, at least in our neighborhood. Our planet is just close enough to the sun to be warm but far enough to not burn. Earth has a large nearby moon which slows down its rotation to lengthen the day and to stabilize it over the long-term. Earth shares the sun with Jupiter, which acts as a comet magnet. The ice of those captured comets may also have given Earth its oceans. Earth has a magnetic core which generates a cosmic ray shield. It has the appropriate level of gravity to retain water and oxygen. It has a thin crust which enables the churn of plate tectonics. Recent research even suggests that there’s a goldilocks zone in the galaxy as well. Too close to the center of the galaxy and a planet is bombarded with constant lethal cosmic radiation; too far from the center and when the planetary mass condenses from star dust it will miss the heavy elements that are needed for life. Such a list can quickly get out of hand to include every aspect of earth. It’s all perfect! It soon resembles one of those phony “Help Wanted” ads engineered to stealthily fit only one favored person.
Some of these factors may indeed be essential for life of any type, but many, if not most, will turn out to be simply coincidental. It will take examination of more planets, and more examples of life to know. But there is an even a stronger assertion about the nature of nature. This claim asserts that not only do the laws of nature shape life, but that, in Paul Davies’ phrase, “the laws of nature are rigged in favor of life.” In this view “life emerges from a soup in the same dependable way that a crystal emerges from a saturated solution, with its final from predetermined by the interatomic forces.” Cyril Ponnamperuma, an early pioneer in biogenesis (study of the origin of life), believed “there are inherent properties in the atoms and molecules which seem to direct the synthesis” toward life. Stewart Kauffman and many others believe, based on their models, that when conditions are right, the emergence of life in inevitable. Mathematician Manfred Eigen wrote in 1971, “The evolution of life, if it is based on a derivable physical principle must be considered an inevitable process.”
Christian De Duve, a Nobel prize winner for his work in biochemistry, believes life is a cosmic imperative. He writes in his book Vital Dust: “Life is the product of deterministic forces. Life was bound to arise under the prevailing conditions, and it will arise similarly wherever and whenever the same conditions obtain… Life and mind emerge not as the result of freakish accidents, but as natural manifestations of matter, written into the fabric of the universe.”
If life is inevitable, why not fishes? If fishes are inevitable, why not mind? Simon Conway Morris speculates that “What was impossible billions of years ago becomes increasingly inevitable.” As the subtitle of his book makes clear, we are “inevitable humans.”
One way to test the cosmic imperative before we make (or don’t make) contact with ET or detect life on a comet, is to rerun the tape of life. (I should finally mention that “rewinding the tape”, like “dialing the phone,” or “cranking the engine,” is a skeuonym, an action continued from a technology no longer practiced; for you young readers, it means to re-run an unfolding sequence from the same starting point.) Gould called rewinding the tape of life the great “undoable” experiment, but he was wrong: it turns out you can.
The new tools of sequencing and genetic cloning make replaying evolution possible. You take a simple bacteria (E. coli), select an individual and make dozens of identical clones of that one particular bug. Genetically sequence the genotype of a clone. Put each remaining clone into exactly identical incubation chambers, with exactly identical settings and inputs. Let the cloned bacteria multiply freely in parallel pots. Let them breed for 10,000 generations. Then 20,000 generations. Let them run for 20 years and you’ll have 40,000 generations and noticeable evolution. At each 1,000 generation milestone, take a few out, freeze them for a snapshot, and sequence their evolved genomes. Compare the parallel evolved genotypes across all the pots. You can re-run the tape of evolution at any time along the way by retrieving a frozen snapshot specimen and redeploying the bug in another identical chamber.
Richard Lenski, at Michigan State University, has been performing this very experiment in his lab. What he has found is that in general, multiple runs of evolution produced similar traits in the phenotype – the outward body of the bacteria and what it does. Changes in the genotype occurred in roughly the same places, though the exact coding was often different. This suggests a convergence of broad form with details left to chance. Lenski is not the only scientist doing experiments like this. Others show similar results from parallel evolution: “the convergence of multiple evolving lines on similar phenotypes.” As geneticist Sean Carroll concludes, “Evolution can and does repeat itself at the levels of structures and patterns, as well as of individual genes… This repetition overthrows the notion that if we rewound and replayed this history of life, all outcomes would be different.” We can rewind the tape of life and when we do, it often turns out roughly the same. The big themes in life seem inevitable, and perhaps in technology as well.
In Gould’s last book before his death, his wordy, 1200-page opus of everything he knew, he began to backtrack a little on his former adamant denial of any direction, or ahistoricity, in evolution. I think several things changed his mind. The correct re-classification of the bizarre Burgess Shale organisms from examples of bizarre unknown phyla to bizarre examples of known phyla and the continued accumulation of “surprising” convergent forms played only a small part. The main impetus for re-casting his view of “the structure of evolutionary theory” (the title of his last book) was new scientific tools and techniques. In the past twenty years genetic analysis became cheap enough that even poor paleontologists and field biologists could sequence the genomes of organisms in great detail. Secondly, cheap, immensely powerful computers arrived on desktops, and the computational perspective of Kauffman’s “order for free” became unavoidable. In these simple simulations, when you ran the tape of life again, you kept finding inherent forms, and new internal evolutionary forces. These illuminated the tilt in evolution. It made it clear that evolution had tendencies, established forms, and a direction.
At the close of his majestic Wonderful Life, Gould imparts his final sermon on greater lessons to be gained from the Burgess Shale – and on this he was not changing his mind. He concluded, “Biology’s most profound insight into human nature, status, and potential lies in the simple phrase, the embodiment of contingency: Homo sapiens is an entity, not a tendency.”
Stephen Jay Gould got it precisely, but elegantly, backwards. If we re-run his sentence again, but this time from back to front, I can’t think of more succinct phrase that sums up evolution’s message better than this:
Homo sapiens is a tendency, not an entity.
Humanity is a process. Always was, always will be. In most respects we have just started our evolution as Homo sapiens. Nothing at all in evolution is fixed. Every organism is on their way to becoming. In evolution all becoming is constrained by past successes, and tilted towards future ones. Much of what happens in evolution is inevitable. We, therefore, are nothing more and nothing less than an ordained becoming.
We are ordained becoming within a framework that is, in Paul Davies’ wonderful phrase, rigged to favor life. Plain logic demands we ask, what rigged it? There is no satisfactory answer to that yet existential question, although there are many candidates. These include: the cosmic evolution of randomly generated rigging rules, or forking multiverses, or subjective anthropomorphic appreciation (it only looks rigged because we are here), or, an eternal favorite, God. Unraveling that quandary of genesis is too large a story to be sorted out here.
Rather than be concerned with the nearly unanswerable quest for the cosmic imperative’s origins, I am far more interested in its destiny. Where is evolution headed ? What technologies are inevitable? And if we are ordained becoming, then what are we ordained to become?
